
Ecology: Biology of Interaction. I-08. Stability of Biosystems
A characteristic feature of regulation by the principle of negative feedback is that it leads to oscillations of the regulated variable. If the influence drives the biosystem beyond the limits of its regulation by negative feedback loops, it will transition to another quality. Positive feedback loops will then become active...
A characteristic feature of regulation by negative feedback is that it leads to oscillations of the regulated variable. It is interesting to compare, for example, the phenomena occurring in a standing table and in a human body standing next to it. The table has undergone elastic deformation, in which the action of gravitational force is balanced by elastic force, and is in a state of rest. A human continuously monitors their posture (using a significant portion of their brain's "capacity" for this). A slight deviation of the body to one side triggers receptors that activate effectors (muscles) that return the system to the required state. The action of these effectors compensates for the deviation with a certain excess. Because of this, the system passes through the optimal state and requires regulatory action in the opposite direction. The body of a standing human is simultaneously involved in several oscillatory processes with different frequencies and amplitudes! This is why a standing person becomes tired and expends energy, while a table neither expends energy nor wears out.
What will happen if a biosystem, whose state is regulated by negative feedback, experiences some external influence that changes its parameters? Let us consider this using the example of population abundance regulation. A population in a stable state maintains its abundance relatively constant, while experiencing continuous fluctuations. Suppose catastrophic events or human impact have destroyed a significant portion of individuals. How will the population respond to this impact?
If the impact drives the population beyond the limits of regulation of its abundance by negative feedbacks, it will transition to another state (for example, it will die; Fig. I-8.1). Thus, if the number of individuals becomes very low, their reproduction may be disrupted (for example, because partners cannot find each other). Positive feedbacks will be activated: a decrease in population abundance will cause a decrease in the birth of new individuals. As a result, the biosystem will transition to another state (the population will die). However, if the impact turns out to be non-critical, negative feedbacks will be able to return the population abundance to normal. This means that a decrease in abundance will cause a weakening of intraspecific competition, enhanced reproduction, and better survival of offspring.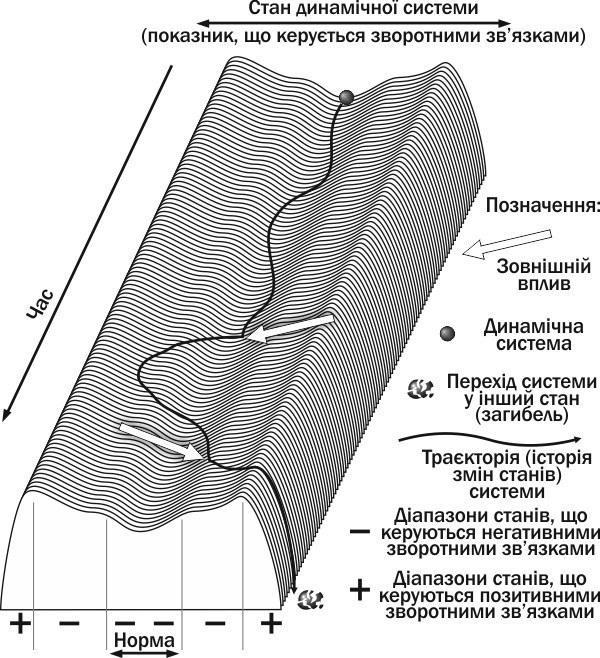
Fig. I-8.1. Trajectory of a system controlled by feedbacks. The first external impact drove the system out of the normal range, but more powerful negative feedbacks returned it to the normal range. The second external impact transferred the system to the range of action of positive feedbacks, which amplified the deviation and led to the system's death
To describe processes similar to those mentioned above (and those shown in Fig. I-8.1), the concept of stability is important. Let us analyze Fig. I-8.1. Is the state of the biosystem in the normal range stable? Yes, and this state is characterized by the fact that deviations from equilibrium trigger mechanisms that return the biosystem back. This stability is not hindered by the fact that the biosystem in question is in a state of continuous changes.
As a result of external impacts, the biosystem whose dynamics are shown in Fig. I-8.1 died. Is the death state stable? Yes, but this is a different stability than in the case of the normal state. The state of a system located on the "ridge" separating the ranges of states controlled by negative and positive feedbacks (as well as a system "rolling down" under the influence of positive feedbacks) can be called unstable. This means that from the standpoint of stability, these states belong to special types.
Thus, for further discussion, it is necessary to define the concept of "stability." It has become one of the most important ecological concepts from the early stages of ecology's development. For example, it was key to F. Clements' climax theory of plant communities that appeared at the beginning of the 20th century.
The stability of a plant community is its ability to "maintain its composition and order for a long time. This stability is determined by the community's ability for self-reproduction" (V. Sukachev).
The use of the concept of "stability" in ecology is a special case of its general scientific application. The classic definition of the concept of "stability" is that given in 1892 by the creator of the theory of motion stability, Alexander Mikhailovich Lyapunov (a docent, and later — a professor of Kharkiv University, now named after V. N. Karazin). Stability was defined by A. M. Lyapunov from the standpoint of the effects of perturbations on motion with certain initial conditions. The application of such an approach to describing biosystem dynamics faces significant difficulties and does not fully correspond to the tradition that has developed in ecology. In our opinion, from mathematical approaches for ecology, the most useful is the concept of invariance by J. L. Lagrange, which is a special case of stability by A. M. Lyapunov. From this standpoint, stability is the ability of a system to remain in a bounded region of its phase space.
Phase space in this case is called the set of possible states of the system, described using a set of its state variables (for example, the movement of the system in phase space is shown in Fig. I-8.1). The choice of variables that define the phase space makes it possible to consider, for example, the stability of a community both from the standpoint of the number of its species and from the standpoint of the invariance of its structure. The essence of the stability phenomenon remains unchanged; only the set of coordinates in which the system trajectory is examined changes.
But what bounds a certain region of the phase space? By the trajectories of systems located in that region. In Fig. I-8.1, one can see certain bounded regions of phase space: the stable equilibrium zone, in which the state of the system is controlled by negative feedbacks, and two zones (to the right and to the left of the stable equilibrium zone), in which the dynamics of the system are determined by positive feedbacks. A special state is, probably, the position on the "ridge" separating the "+" and "—" control regions, as well as the state of the dead system. These regions of phase space can be called stability basins. Each stability basin corresponds to its own dynamic type of the considered biosystems (the concept of dynamic type is analyzed in more detail in paragraph I-12).
Based on the results of studying changes in the composition of hemiclonal population systems of the green frog hybrid complex, the authors of this manual have proposed a classification of types of biosystem stability, shown (using physical analogies) in Fig. I-8.2. It can be assumed that this classification is applicable not only to the studied category of systems, but also to any others.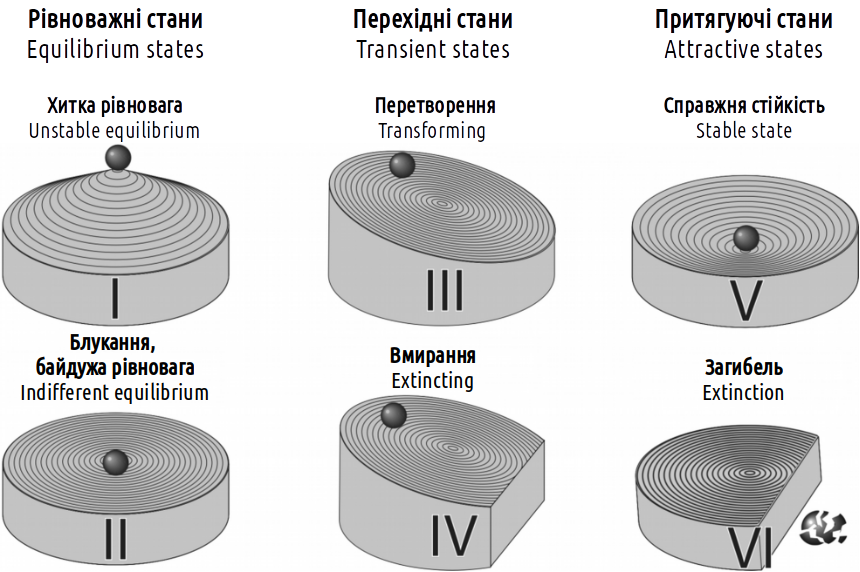
Fig. I-8.2. Classification of types of system stability
The difference between the physical analogies used in Fig. I-8.1 and I-8.2 is related to the fact that in the first figure, the character of the surface profile (which determines the dynamics characteristic of a certain type of stability) is shown in one plane, and in the second — in two perpendicular planes. Analyzing the dynamics of the system in Fig. I-8.1, one can find all types of stability shown in Fig. I-8.2, except for type II (wandering). This case corresponds to a situation when a change in the system state turns out to be indifferent to the mechanisms that govern its dynamics. Type I (precarious equilibrium) corresponds to the position on the "ridge" separating the regulation ranges of negative and positive feedbacks. A deviation of the system from the state of precarious equilibrium will lead to its transition to one of the alternative states.
It should be emphasized that in most cases, authors writing about system stability mean precisely the states of their genuine stability (type V according to the given classification). Naturally, in many cases, such states are of the greatest interest. For example, the task of protecting a particular biosystem (for example, a valuable ecosystem in a protected territory) can be presented as the task of maintaining it in type V stability. A characteristic feature of this state is that in case of deviation from the equilibrium state, a force arises that returns the system to the same equilibrium state.
The dynamics of changes in the parameters of a system in a state of type IV stability is of significant interest for its diagnosis (and prediction of its future changes). How quickly a changed system will return to its normal state is an important characteristic of it. Analyzing its dynamics, two relatively independent parameters can be distinguished: the ability to resist impact and the ability to return to normal after change. These parameters reflect different properties of the system (they can be compared to hardness and elasticity in mechanics: a diamond is hard but not elastic, and rubber is not hard but elastic). These parameters are negatively correlated (as are strength and elasticity in classical mechanics). They are two components of overall stability (shown in Fig. I-8.3 as the area between the curve describing the dynamics of the system state and the "corridor" of normality).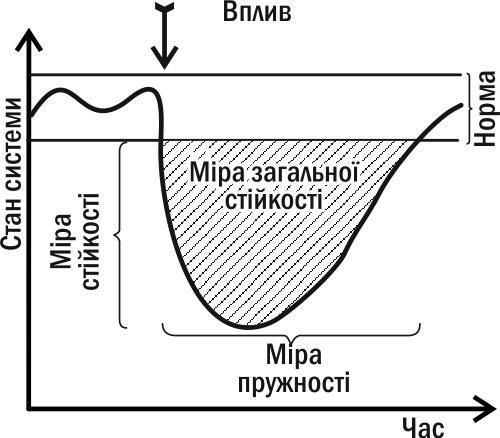
Fig. I-8.3. Main measures of stability applicable to biosystems in a state of type IV stability (see Fig. I-8.2)