
Ecology: biology of interactions. III-08. Succession. Basic concepts
The main cause of successions is the absence of equilibrium between production and respiration in the ecosystem, that is, an imbalance of ecological balance. This leads to a change in the stock of organic matter in the ecosystem and, ultimately, to a change in the community. Placing various ecosystems on the plane "production-respiration"...
III-8. Successions. Basic Concepts
Have you ever found yourself on an abandoned field or in an abandoned garden? You probably realize that by their outward appearance one can determine when they were last tended. How is this possible? An ecosystem left to itself (in these examples—an artificial agricultural ecosystem—agroecosystem) begins to change. Knowing the rate of such changes, one can estimate how long ago it was abandoned.
Now imagine a river flowing through a wide valley. The channel of such a river forms loops—meanders. Where the channel turns, the flowing water erodes the bank it meets and forms a bank cliff. And on the opposite side of such a bank, a sandy beach grows. Near the water the sand is bare, a little farther—it is overgrown with grass, still farther—the grass forms a dense sod, and finally, at some distance from the bank, shrubs grow. It is easy to understand that this transition from one community to another (ecocline) reflects the dynamics of changes in each individual section of the bank over time. Where shrubs grow now, once only a beach was forming, which once created a new section of the bank... The loops of the river grow and eventually connect with each other. The channel finds a shorter path, and its former sections form oxbow lakes. Eventually, the oxbow lakes transform into floodplain lakes, fill with sediments, and become parts of a floodplain meadow. The examples we have given demonstrate the ability of ecosystems to change over time—successions (from Lat. successio—inheritance, succession).
Succession is the sequential change of communities in one habitat. This is a directed process, controlled by the community, leading to a certain state. Succession in biogeocenoses is a more prolonged process than seasonal changes, but not as prolonged as the evolution of ecosystems. An ecocline, thus, is a succession observed in space.
Usually in the course of succession, transient stages can be distinguished, called serial communities (or simply series), and the final stable state—the climax community (climax). The theory of successions was developed in the 1920s by Frederick Clements, an American ecologist, who regarded succession as a kind of analog of ecosystem ontogeny.
The cause of successions is not only the change of the environment by some external factors (for example, flowing water, as in the described case with the river meander), but also the entire set of interactions between components of communities, and sometimes—the influence of humans.
"The characteristic dynamics of communities described for the end of the 19th—beginning of the 20th centuries in the Scottish heath bogs. The owner of these lands used them for hunting the numerous Scottish grouse found there. In 1892–1893, several pairs of gulls settled on the heath. Protected by the owner, at the beginning of our century they already formed a large colony numbering 1,500–2,000 pairs. The birds abundantly fertilized the soil; the hygroscopic material of their nests accumulated moisture. As a result, gradual paludification began, which led to the gradual disappearance of heather and its replacement by harsh grasses. The lands began to be used as hay meadows. However, later the grasses were displaced by rushes, and later—by sorrel and other weeds. Simultaneously, the number of grouse decreased; on the waterlogged areas ducks appeared—a species less valuable as a hunting trophy. All this led to the cancellation of gull protection; moreover, their nests were deliberately destroyed. In 1917, about 30 pairs remained, the heath was gradually restored, the ducks disappeared, and the grouse reappeared. However, even to this day, the restoration of the original community has been incomplete: on the heath waste, grasses and some weeds have remained—a 'trace' of the successional series" (I.O. Shilov, 1998).
The main cause of successions is the absence of balance between production and respiration in the ecosystem, i.e., imbalance of ecological balance (see section III-5). This leads to a change in the store of organic matter in the ecosystem and, ultimately, to a change in the community. Placing various ecosystems on the 'production-respiration' plane (Fig. III-8.1), we can see that only those in which production and respiration balance each other prove to be stable.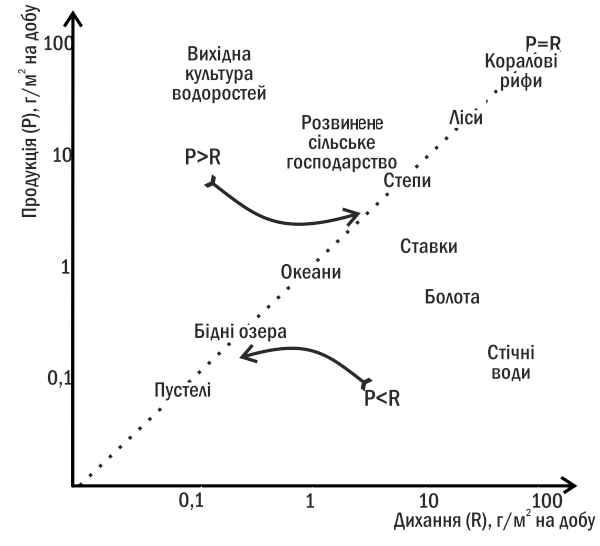
Fig. III-8.1. Ordination (placement) of different types of ecosystems on the 'production - respiration' plane
If in some community production exceeds respiration (for example, in the initial culture of algae—a solution of mineral salts populated by phytoplanktonic organisms), autotrophic succession begins in it. An excess of organic matter accumulates in such an ecosystem, changing its properties and creating an environment for the reproduction of heterotrophs. Production in the community decreases, respiration increases, and eventually these two values become equal.
Heterotrophic succession (Fig. III-8.2) begins from a state in which respiration of the community exceeds production. Gradually, heterotrophs destroy the excess of organic matter, and equilibrium between respiration and production is achieved in the system.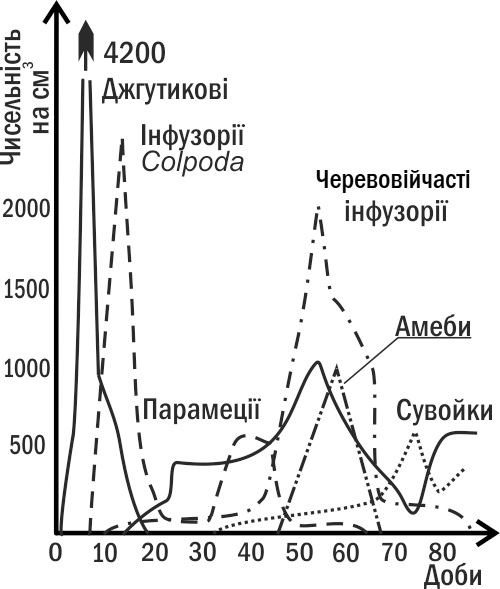
Fig. III-8.2. Example of heterotrophic succession: dynamics of numbers of different protozoa in hay infusion
A characteristic case of autotrophic succession can be considered the colonization of a territory by organisms where there are no reserves of organic matter. Let us consider the following example. Lichens settle on bare rock, gradually corroding the stone with acids they secrete. Soil appears in the cracks of the rocks. Mosses begin to grow on the rock, and then grasses. Forming a sod of interwoven dead stems and rhizomes, grasses retain detritus and even collect dust. After a very long time, shrubs appear on the site of the former rocks, and then a forest. This was primary succession—a succession that occurred where there were no reserves of organic matter from former communities. Another example of primary succession is shown in Fig. III-8.3.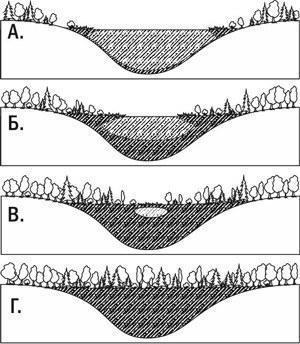
Fig. III-8.3. Example of primary succession: siltation of a lake
After some time, the forest that grew during primary succession is destroyed by fire. A burn forms. On the burn, remains of forest soil (thickly sprinkled with ash) and even seeds of many plant species are preserved. Already the next year, the burn becomes overgrown with grasses (for example, fireweed is very characteristic in such conditions). After a few years, shrubs and young trees will appear on the site of the burn, and after a few decades or a century, a forest will develop here, resembling the one that grew here before the fire. Secondary succession occurs in habitats where remains of organic matter from previous stages of development of this ecosystem have been preserved (Fig. III-8.4.)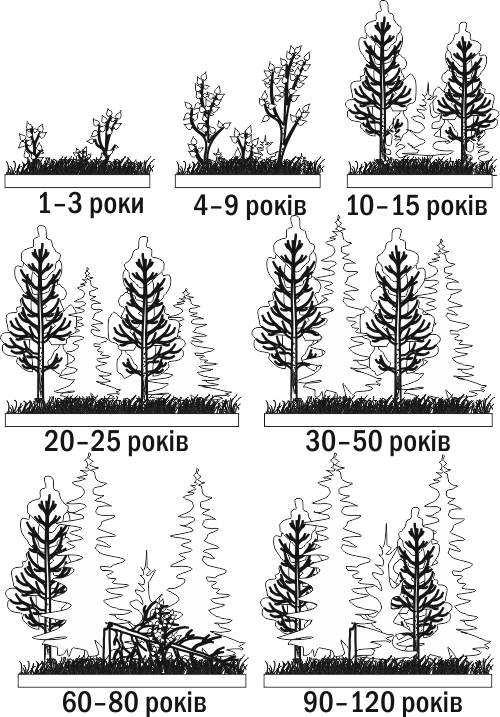
Fig. III-8.4. Example of secondary succession: overgrowing of a burn by mixed forest
Heterotrophic succession requires a reserve of organic matter for its course. Most examples of heterotrophic succession that we can observe unfold not on the scale of biogeocenoses, but within relatively small ecosystems. For example, these include the bodies of dead animals or trunks of fallen trees. Successions occurring in such temporary ecosystems are called destructive—they do not lead to climax, but simply destroy the temporary habitat in which they occur. Sometimes ecoclines can also be observed in heterotrophic successions. Dig up fallen pine needles and the upper layer of soil in a coniferous forest. From above, the needles are almost undamaged, but the deeper—the more they are transformed by bacteria, fungi, and detritivore animals. Each fallen needle undergoes this path of destruction, corresponding to the sequence of layers of coniferous litter.
Clements was a proponent of the monoclimax concept. Currently, the polyclimax concept has prevailed, according to which for each region there is a certain climatic or zonal climax, and, in addition, certain edaphic (i.e., determined by soil characteristics) or local climaxes can be stable within it. Cyclical climax has also been described, in which the ecosystem passes through a set of certain states that change one another in a circle. Sometimes these changes are determined by catastrophic events that regularly occur at a certain stage of ecosystem development. Thus, a spruce forest after reaching a certain maturity can be regularly destroyed by pests; chaparral can regularly burn down in fires.
One can consider both general and special successions, concerning changes in species in certain ecological niches. Thus, in an orchard, young trees are fed upon by polyphagous phytophagous insects: beetles, weevills,geometers,silk worms, tortricids. When the trees begin to bear fruit, fruit moths, blossom weevils, and curculios appear, feeding on flowers and fruits. As the orchard ages, trunk pests inhabit it (bark beetles, longhorn beetles, clearwing moths).