
Ecology: the Biology of Interaction. V-13. Heat Balance and Thermobiological Types of Organisms
Appendices: Syllabus. Questions. References. Personalities. Glossary. R Commands.
V-13. Heat Balance and Thermobiological Types of Organisms
In cases where some parameter important to biosystems is influenced by a variety of processes, it is customary to treat the regulation of that parameter as a balance. As we have already noted, temperature is the most critical condition, exerting a profound influence on the course of all biological processes. The body temperature of any organism is affected by a whole range of factors (Fig. V-13.1).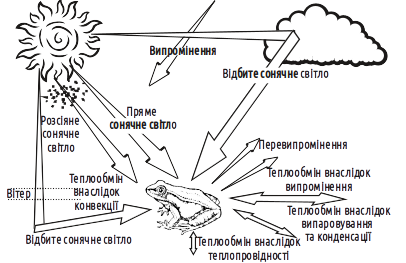
Fig. V-13.1. The heat balance of an organism depends on many factors
Examining the adaptations of organisms for maintaining their body temperature, we can see how all the features of their way of life are reflected in these adaptations. For instance, organisms that maintain a constant body temperature exceeding that of the surrounding environment (homeothermic organisms) possess a variety of adaptations for heat conservation. In many mammals this is fur; in birds, feathers. The characteristic body shape of whales and seals is determined not only by the refinement of their streamlining but also by the thick adipose “jacket” enveloping their bodies. If certain body parts of homeothermic organisms are subjected to intense cooling (such as the legs of gulls, with which they can walk on ice), specialised heat-conserving structures may develop in them. Thus, the blood vessels in the legs of a gull operate on the countercurrent principle. Arteries carrying warm blood to the legs and veins carrying cold blood away from them lie in close proximity. Heat is transferred from the arterial blood to the venous blood. If the vessels were not in close apposition, there would be no effective heat exchange between them. And if the blood flows were directed in the same direction, the temperatures of the arterial and venous blood would simply be averaged (and for the arterial blood entering the foot would be approximately 16°C, which would result in substantial heat loss through the legs). Thanks to the countercurrent arrangement, a large portion of the heat is released by the arterial blood before it enters the legs and is returned to the trunk via the venous blood.
The smaller the body size of an animal, the more difficult it is for it to maintain a constant body temperature. The minimum body size of homeothermic animals presumably corresponds to that of shrews and hummingbirds. The larger the animal, the easier it is for it to maintain a constant body temperature. For large animals the principal challenge becomes the risk of overheating (for example after a period of intense muscular activity). It is no coincidence that large homeothermic animals living in warm climates generally do not have a dense fur coat, and may even possess specialised “radiator” organs that serve to dissipate excess heat. The ears of elephants perform precisely this function; when overheated, elephants increase blood circulation in their ears and fan them.
An effective means of reducing body temperature is the evaporation of water from the body surface. This water may evaporate from specific areas of the body (from the oral cavity, as in dogs or crocodiles) or from the greater part of the body surface (as in humans). The mode of heat balance regulation in humans is, in general, rather unusual. Our species evolved as a species of non-specialised hunters and gatherers of the African savanna. One of the significant advantages of our ancestors was their high endurance. It may be difficult for the pampered inhabitants of cities to believe, but a trained human can run longer and cover a greater distance than a trained horse! During such prolonged running, the muscles generate a considerable quantity of heat. Probably the only means of cooling under such conditions is intense sweating. Humans possess an extraordinarily high density of sweat glands per unit of body surface. Were they covered with a dense coat of hair, as our closest relatives are, the settling dust would form a crust over the body. The loss of fur in our case is an adaptation for “shedding” excess heat. However, our ancestors were required not only to dissipate surplus heat into the environment but also to conserve it, for instance during cool nights. This is associated with the development in humans of a fairly substantial layer of subcutaneous fat, considerably greater than in the great apes.
There are a considerable number of ways of regulating the heat balance, but they can be classified by division into several groups. The most “obvious” division is that of organisms into poikilothermic and homeothermic. These two groups may carry other names. The concepts of “cold-blooded versus warm-blooded” and “ectothermic versus endothermic” should be considered unsatisfactory and their use avoided. These terms refer to features of organismal thermoregulation that should not be regarded as primary. In cold-blooded organisms the blood may be quite warm, and endotherms may derive a substantial share of their energy from external sources. The terms “poikilothermic” (Greek poikilos — variegated, diverse, and therme — heat, warmth) and “homeothermic” (Greek homoios — similar, uniform) are free of these shortcomings and refer to the primary characteristic: variable or constant body temperature.
Examining these groups of organisms, one can observe that transitions exist between them. Let us consider a more detailed classification (Fig. V-13.2). By type of thermoregulation, organisms are divided into the following thermobiological types:
— poikilothermic (which do not maintain a constant body temperature through the operation of specialised physiological systems):
— aregulatory poikilothermic (capable of temperature regulation only through selection of the most favourable environmental conditions);
— regulatory poikilothermic (capable of temperature regulation through the presence of mechanisms for warming or cooling the body);
— gigantothermic (possessing a relatively constant body temperature maintained by virtue of large body size);
— homeothermic (which maintain both the upper and lower limits of the range of normal temperatures):
— heterothermic (capable of maintaining a constant body temperature, as well as substantially lowering it when necessary).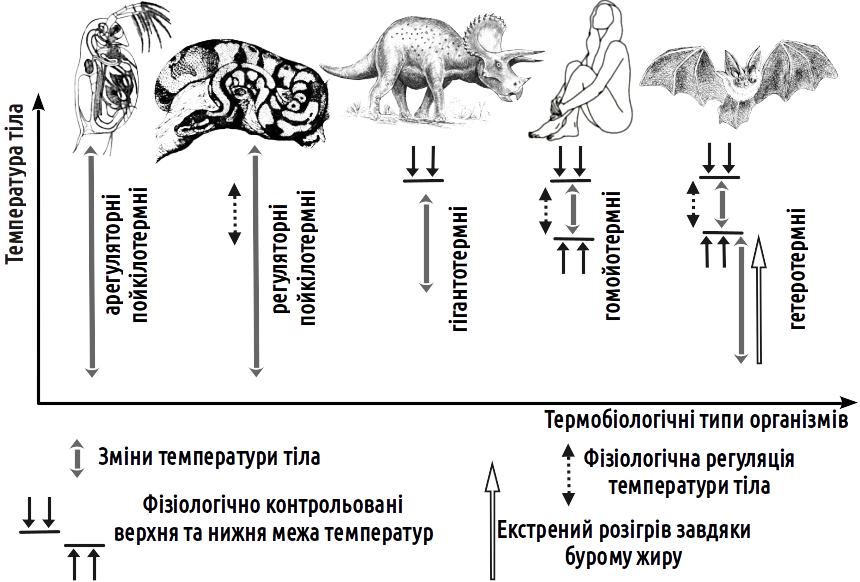
V-13.2. Body temperature regulation in representatives of different thermobiological types of organisms
Why do regulatory poikilotherms not maintain a constant body temperature? To conserve energy under conditions in which this is not necessary. Nevertheless, during rapid swimming, tunas and opah (Lampris guttatus, Fig. V-13.3) maintain an elevated body temperature. Hawk-moths warm up before flight, and pythons warm their egg clutches.
Among homeothermic animals, the majority of birds and mammals are included; beyond these, homeothermy arose in flying reptiles (pterosaurs) and small predatory dinosaurs (presumably closely related to birds). Conversely, certain mammal species are not truly homeothermic, such as the naked mole-rat Heterocephalus glaber (an African colonial rodent with a social organisation resembling that of social insects, Fig. V-13.3).
Heterothermy may be seasonal in character (hedgehogs, ground squirrels, marmots) or even diurnal (bats and hummingbirds). In order to rewarm, heterothermic organisms must possess an alternative heating system.
For example, if the body temperature of a human falls below 35°C, the rate of their metabolic processes also declines sharply. Under such conditions the individual cannot rewarm spontaneously and return to a normal state (however, if such a hypothermic person is rewarmed using an external heat source, they can be restored to a normal state). In heterothermic animals, a specific tissue (brown adipose tissue), capable of rapid generation of large quantities of heat through peroxisomal oxidation of lipids, serves for the rapid elevation of body temperature following cooling.
The thermoregulation of dinosaurs is one of the questions debated with considerable intensity in the popular literature. Curiously, although direct investigation of this problem is no longer possible, we know rather more about it than might be supposed.
Large dinosaurs were gigantothermic animals. As calculations demonstrate, in a subtropical climate an animal with a mean body diameter of 1 m (and many dinosaurs were larger) would have a body temperature of 34°C with diurnal fluctuations of less than 1°C, without any additional energy expenditure by the animal itself. The largest dinosaurs were in no danger of becoming hypothermic, since they were truly large. Overheating was the more likely threat to them. In this regard, our reconstructions of dinosaurs suffer from a serious deficiency: we do not depict on them the organs that might have performed the function of dissipating excess heat (as the ears of the elephant or the moist tongue of the dog do).
Gigantothermy may have represented a transition from poikilothermy to homeothermy. The enzymes of gigantothermic animals were specialised for functioning at a constantly elevated body temperature. At the same time, physiological mechanisms developed in such organisms that permitted control of the upper thermal limit, thereby avoiding overheating. In practical terms, gigantotherms enjoyed the advantages of homeothermy without expending energy to achieve it! What might have “compelled” gigantotherms to begin maintaining a constant body temperature, incurring energetic costs in doing so?
Homeothermy arose several times in the history of life. According to one point of view, in the majority of instances its emergence proved to be associated with a reduction in body size. Thus, in mastering flight, the ancestors of birds and pterosaurs reduced their body size (it will be recalled that flight is easier for a smaller animal). The ancestors of mammals reduced their size after losing in competition with dinosaurs in the larger size class (sometimes a tactical defeat leads to a strategic victory). Crocodiles, however, for example, although they also reduced their body size, did so in the course of adaptation to lying in ambush at the bottom of bodies of water, which did not permit energy expenditure on thermoregulation. As a consequence, crocodiles remained poikilothermic.
Do gigantothermic animals exist today? To some degree, elements of gigantothermy can be identified in the heat balance of large lizards, snakes, crocodiles, and tortoises. Of particular interest is the mode of thermoregulation of the largest extant tortoise species, the leatherback sea turtle Dermochelys coriacea (Fig. V-13.3).
“A curious peculiarity of this genus is its ability to maintain a body temperature at least 18°C above that of the surrounding water. … As in mammals and birds, this is provided by muscular activity and is sustained by an external insulating layer of fat and countercurrent heat exchange in the blood vessels of the limbs” (Robert Carroll, 1992).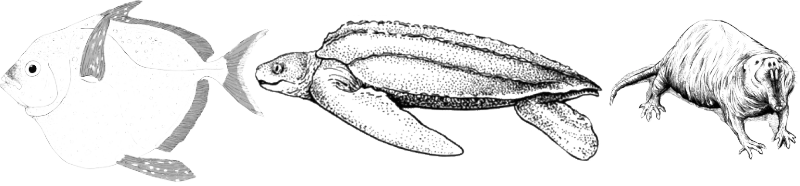
Fig. V-13.3. Some animals exhibit modes of thermoregulation different from those that might be expected on the basis of their systematic position. The opah, the leatherback sea turtle, and the naked mole-rat demonstrate this point
In all probability, the leatherback sea turtle can be regarded as the animal most closely resembling medium-sized dinosaurs in its thermoregulation. Large dinosaurs had no need to conserve heat, while small ones were homeothermic!