
Ecology: Biology of Interactions. III-13. Ecological Pyramids
Charles Elton proposed a way of graphically expressing relationships between trophic levels, which has become almost a symbol of ecology as a science. This refers to ecological pyramids. When constructing ecological pyramids, measures of abundance of representatives of different trophic levels are shown as rectangles, st...
III-13. Ecological Pyramids
Charles Elton proposed a way of graphically expressing relationships between trophic levels, which has become almost a symbol of ecology as a science. This refers to ecological pyramids. When constructing ecological pyramids, measures of abundance of representatives of different trophic levels are shown as rectangles stacked on top of one another. This method is typically used to describe grazing trophic chains. Pyramids of numbers, biomass, and productivity are distinguished.
Let us also construct several ecological pyramids. Our examples will be fairly simplified: we will assume that the trophic chains we describe have no 'branches.' For example, modeling the chain 'clover - sheep - wolves,' we will assume that sheep feed only on clover and wolves feed only on sheep, and we will be interested in the relationships between these trophic levels within some ecosystem where the abundance ratios of the considered levels have reached equilibrium. Naturally, ecological pyramids can also be used to describe natural ecosystems, and the simplifications we adopt are needed only to make our reasoning more illustrative.
Let us return to the chain 'clover - sheep - wolves.' Evaluating the population sizes in this chain, we will see that individual clover plants far outnumber sheep, and sheep outnumber wolves. Such a pyramid (with a base wider than the apex) is called a regular or upright pyramid. However, not all grazing chains produce upright pyramids of numbers. An example is the chain 'oaks - oak silkworms.' On a small number of large trees, many caterpillars can live. The pyramid of numbers for this chain will be inverted (Fig. III-13.1).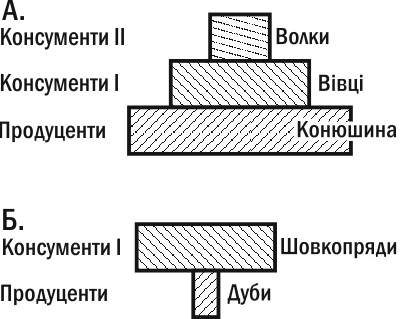
Fig. III-13.1. Pyramids of numbers. A. Upright. B. Inverted. The inverted nature of the second pyramid is associated with differences in individual sizes.
It is easy to understand that the inversion of the second pyramid is associated with differences in the sizes of producers and consumers. Based on data about the average weight of an oak and an average silkworm, as well as the pyramid of their numbers, we can construct a biomass pyramid. Naturally, it will be upright (Fig. III-13.2).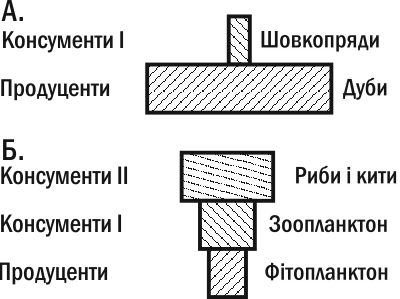
Fig. III-13.2. Biomass pyramids. A. Upright. B. Inverted. The inverted nature of the second pyramid is associated with differences in 'speed of life' among individuals -- the varying intensity of energy flow through their biomass.
Can biomass pyramids be inverted? Quite rarely, but they can. Let us consider the grazing trophic chains of the ocean water column. Surprisingly, the biomass of producers (planktonic algae) in such chains is often less than the biomass of consumers. Does this mean that consumers in such chains exist independently of producers? No.
A simple analogy will help clarify this. A large pond can exist thanks to a small stream flowing into it, even though at any given moment the mass of water in the pond is far greater than in the stream. This is clearly possible because the water in the stream is renewed much more rapidly than in the pond. Similarly, in water-column communities, energy flows through different trophic levels at different rates. The turnover time of phytoplankton biomass is measured in hours, zooplankton in days, fish and whales in weeks and months. To account for this difference, we need to reflect the intensity of energy flow through each level in ecological pyramids. Based on data on the biomass of trophic chain links and the rate of its change, we can construct a pyramid of productivities (or energy flows; Fig. III-13.3).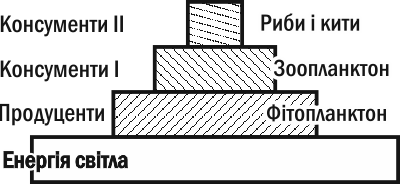
Fig. III-13.3. Productivity pyramids are always upright.
Such a pyramid will always be upright. The first law of thermodynamics (conservation of energy) 'prohibits' such a pyramid from being inverted, and the second law prohibits it from having 'floors' of equal width, since with each energy transformation part of it must be dissipated as heat. This is why real trophic chains are never very long, and ecological pyramids are never very tall. In any real ecosystem, a consumer at trophic level X would receive so little energy (after ten successive transformations!) that it would be impossible to collect the required energy from the territory accessible to a single individual.
Now that we have familiarized ourselves with the logic behind constructing ecological pyramids, let us consider two more specific examples. Eugene Odum calculated the parameters of a hypothetical food chain in which a twelve-year-old boy subsisted exclusively on veal (note: eating only meat is unnatural!), and the calves fed only on alfalfa (this is more physiologically plausible, ignoring the fact that both the boy and the calves need to begin life feeding on their mothers' milk). The characteristics of such a pyramid are given in Table III-13.1.
Table III-13.1. An example of ecological pyramids for a hypothetical trophic chain
|
Such a pyramid is always upright. The first law of thermodynamics (energy conservation) “forbids” this pyramid from being inverted, and the second law forbids levels of equal width, because at every energy transformation some energy must be dissipated as heat. This is also why real trophic chains are not very long and ecological pyramids are not very tall. In any real ecosystem, so little energy would reach a level-X consumer (after ten consecutive transformations) that the area available to one individual could not provide the required amount of energy. |
Now that we have understood the logic of ecological pyramids, let us examine two more concrete examples. Eugene Odum calculated parameters of a hypothetical food chain in which a 12-year-old boy fed exclusively on veal (note: eating only meat is physiologically unnatural), and calves fed only on alfalfa (more physiological, except that both the boy and calves begin life feeding on their mothers’ milk). The characteristics of this pyramid are shown in Table 3.13.1. |
Table 3.13.1. Example of ecological pyramids for a hypothetical trophic chain |
|
|
Numbers |
1 |
Productivity |
8,3×103 |
|
1 |
4,5 |
8.3×10^3 |
1,2×106 |
|
4.5 |
2×107 |
1.2×10^6 |
1,5×107 |
|
2×10^7 |
— |
— |
6,3×109 |
III-13. Ecological Pyramids
Charles Elton proposed a way of graphically expressing relationships between trophic levels, which has become almost a symbol of ecology as a science. This refers to ecological pyramids. When constructing ecological pyramids, measures of abundance of representatives of different trophic levels are shown as rectangles stacked on top of one another. This method is typically used to describe grazing trophic chains. Pyramids of numbers, biomass, and productivity are distinguished.
Let us also construct several ecological pyramids. Our examples will be fairly simplified: we will assume that the trophic chains we describe have no 'branches.' For example, modeling the chain 'clover - sheep - wolves,' we will assume that sheep feed only on clover and wolves feed only on sheep, and we will be interested in the relationships between these trophic levels within some ecosystem where the abundance ratios of the considered levels have reached equilibrium. Naturally, ecological pyramids can also be used to describe natural ecosystems, and the simplifications we adopt are needed only to make our reasoning more illustrative.
Let us return to the chain 'clover - sheep - wolves.' Evaluating the population sizes in this chain, we will see that individual clover plants far outnumber sheep, and sheep outnumber wolves. Such a pyramid (with a base wider than the apex) is called a regular or upright pyramid. However, not all grazing chains produce upright pyramids of numbers. An example is the chain 'oaks - oak silkworms.' On a small number of large trees, many caterpillars can live. The pyramid of numbers for this chain will be inverted (Fig. III-13.1).
Fig. III-13.1. Pyramids of numbers. A. Upright. B. Inverted. The inverted nature of the second pyramid is associated with differences in individual sizes.
It is easy to understand that the inversion of the second pyramid is associated with differences in the sizes of producers and consumers. Based on data about the average weight of an oak and an average silkworm, as well as the pyramid of their numbers, we can construct a biomass pyramid. Naturally, it will be upright (Fig. III-13.2).
Fig. III-13.2. Biomass pyramids. A. Upright. B. Inverted. The inverted nature of the second pyramid is associated with differences in 'speed of life' among individuals -- the varying intensity of energy flow through their biomass.
Can biomass pyramids be inverted? Quite rarely, but they can. Let us consider the grazing trophic chains of the ocean water column. Surprisingly, the biomass of producers (planktonic algae) in such chains is often less than the biomass of consumers. Does this mean that consumers in such chains exist independently of producers? No.
A simple analogy will help clarify this. A large pond can exist thanks to a small stream flowing into it, even though at any given moment the mass of water in the pond is far greater than in the stream. This is clearly possible because the water in the stream is renewed much more rapidly than in the pond. Similarly, in water-column communities, energy flows through different trophic levels at different rates. The turnover time of phytoplankton biomass is measured in hours, zooplankton in days, fish and whales in weeks and months. To account for this difference, we need to reflect the intensity of energy flow through each level in ecological pyramids. Based on data on the biomass of trophic chain links and the rate of its change, we can construct a pyramid of productivities (or energy flows; Fig. III-13.3).
Fig. III-13.3. Productivity pyramids are always upright.
Such a pyramid will always be upright. The first law of thermodynamics (conservation of energy) 'prohibits' such a pyramid from being inverted, and the second law prohibits it from having 'floors' of equal width, since with each energy transformation part of it must be dissipated as heat. This is why real trophic chains are never very long, and ecological pyramids are never very tall. In any real ecosystem, a consumer at trophic level X would receive so little energy (after ten successive transformations!) that it would be impossible to collect the required energy from the territory accessible to a single individual.
Now that we have familiarized ourselves with the logic behind constructing ecological pyramids, let us consider two more specific examples. Eugene Odum calculated the parameters of a hypothetical food chain in which a twelve-year-old boy subsisted exclusively on veal (note: eating only meat is unnatural!), and the calves fed only on alfalfa (this is more physiologically plausible, ignoring the fact that both the boy and the calves need to begin life feeding on their mothers' milk). The characteristics of such a pyramid are given in Table III-13.1.
Table III-13.1. An example of ecological pyramids for a hypothetical trophic chain
|
|
Fig. 3.13.4. Biomass of several mammal species in a North American deciduous forest |
As you can see, ecological pyramids cannot be very tall because part of the energy is lost when moving from one level to another. However, different organisms lose different amounts of energy. In different communities, the average level of ecological efficiency differs and is closely related to the number of trophic levels, as shown in Table 3.13.2. |
|
Table 3.13.2. Mean number of trophic levels in different biomes (R. Ricklefs, 1977) |
25% |
7,1 |
|
Average number of trophic levels |
20% |
5,1 |
|
7.1 |
10% |
4,3 |
|
5.1 |
5% |
3,2 |
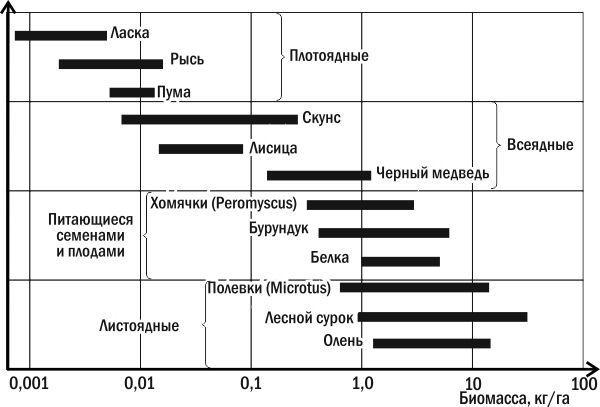
Of course, ecological efficiency varies greatly among different trophic levels, and it is especially low at the base of ecological pyramids. Feeding on plant food is a more complex biochemical and physiological 'task' than feeding on animal food. In most terrestrial ecosystems, a surplus of plant food is registered. However, the number of herbivorous animals (first-level consumers) is typically well controlled by carnivores. A surplus of organisms at this level will effectively be 'consumed' by organisms of the next level.
According to the natural equilibrium hypothesis, the primary regulator of the balance between trophic levels is the apex predator -- the consumer at the highest level. Thus, in a system with an even number of trophic levels, odd levels (1st, 3rd, 5th ...) are more effectively controlled by consumers, while in a system with an odd number of levels, even levels are. Since in terrestrial ecosystems the most difficult level to control is the first -- the producer level -- one would expect an odd number of trophic levels to be more commonly observed in terrestrial systems. Observations confirm this hypothesis.