
Ecology: The Biology of Interaction. IV-10. Predation
An important characteristic of a polyphagous predator capable of switching from one prey to another is its response to changes in relative prey density. A predator with a Type I response selectively consumes a particular prey species, even when they are rare, and increases their consumption as their prevalence grows. A predator with a Type II response...
IV-10. Predation
The most important characteristic of predators is how specialized they are in feeding on particular prey types. For example, the common fox is an extremely plastic predator, whose prey can include a wide variety of small and even medium-sized mammals, birds, reptiles, amphibians, various invertebrates, and many different plants. Depending on the food types available in its environment, a fox may feed on turtles as well as grapes.
At the opposite pole of highly specialized predators are, for instance, sea snakes of the genus Microcephalophis, Microcephal (literally “small‑headed”). The head and anterior part of the body of this medium‑sized snake are very narrow, "calibrated" to the size of the burrows of benthic sea eels, which these snakes feed on. Another example of high specialization are koalas, arboreal representatives of Australian marsupials. They feed only on young eucalyptus leaves.
Thus, based on the degree of specialization in feeding on certain prey categories, predators can be divided into generalists, like the fox, and specialists, like the microcephalids. A related classification divides predators into monophages (adapted to feeding on a single prey type), oligophages (oriented toward several closely related prey categories) and polyphages (capable of feeding on quite diverse prey). These classifications are not entirely identical, although specialized predators are usually oligophages or monophages, and generalists are polyphages. However, sometimes a relatively narrow adaptation helps to exploit a group of prey with similar traits. For example, the giant anteater is highly specialized in feeding on ants and termites: its claws allow it to break into termite mounds and reach the ants, and its long sticky tongue, protruding from a tube‑shaped, toothless snout, enables it to grasp and swallow hundreds of insects at once. Apart from ants and termites, the anteater cannot feed on anything else, yet virtually any of the numerous species of these social insects can become prey for this oligophagous predator.
A specialized predator can be more efficient than a non‑specialized one, allowing it to partially avoid competition. Conversely, polyphages may have a more balanced diet, find food more easily, and maintain a more stable population size.
Predation is one of the most important mechanisms of energy transfer from one population to another and therefore a key process that makes an ecosystem a cohesive whole. Below are some examples of predator impacts on the species diversity of their prey.
On the littoral (coastal seabed) zones in New England (USA) there are alternating patches covered with hard red algae of the genus Chondrus and patches dominated by soft green algae Enteromorpha. Researchers sought to determine which factors dictate which algae dominate a given patch: whether it becomes covered with Chondrus or Enteromorpha. It turned out that the main consumer of the algae is the periwinkle snail Littorina. Littorina prefers Enteromorpha and avoids Chondrus. When adult periwinkles are transplanted onto patches covered with Enteromorpha, they graze the green algae, and the freed space is colonized by red algae. If periwinkles are removed from a red‑algae patch, the Chondrus surface becomes overgrown with Enteromorpha, which later overtakes the area. Thus, Chondrus grows where periwinkles are present, and Enteromorpha where they are absent. But why are the snails present in some places and absent in others?
Littorina larvae have a planktonic lifestyle. Floating larvae settle on the shore everywhere – both in Chondrus mats and in Enteromorpha mats. In Chondrus mats the larvae develop normally, whereas on Enteromorpha patches they are quickly eaten by shore crabs (Carcinus).
Hence, red algae grow where crabs are absent, and green algae where crabs are present. What causes this distribution? Why don’t crabs live everywhere?
Crab distribution is limited by predation from gulls. Gulls fly over the entire coastline, and crabs hide from them beneath dense Enteromorpha cover. Therefore, where Enteromorpha grows, crabs can hide from gulls and consume the snails that would otherwise cause the decline of Enteromorpha… Inter‑species diversity of coastal waters is maintained at a high level by interspecific links based on predation!
In general, a situation in which a predator prevents the decline of its prey’s species diversity is quite typical. In steppe reserves of the former USSR, herds of ungulates – wild horses (tarpans), wild donkeys and further south even onagers and saigas – once grazed. By feeding on fast‑growing plants, these ungulates prevented slow‑growing species from being displaced. After humans exterminated the wild ungulate herds, domestic herds of cattle and sheep were introduced. When these steppes became protected areas, the cessation of grazing led to the disappearance of rare grass species. The situation can be remedied by moderate grazing – for example, by horses.
After the introduction of rabbit myxomatosis to the British Isles in 1964, the numbers of these herbivores sharply declined. As a result, meadow plant diversity decreased. Rabbits are non‑specialized, almost like mowers, and they graze all meadow plants in succession, thereby preventing more competitive plants from displacing weaker and rarer ones.
Usually polyphagous predators most intensively exploit the most abundant of their potential prey, thereby equalizing their relative abundances.
Have you ever spent a long time collecting mushrooms or berries or fishing with a rod? Recall what you saw after closing your eyes. Leaves and berries, mushroom‑covered fir branches or a bobbing float on the waves – depending on what your attention was directed toward. Depending on the activity you were engaged in, a certain search pattern was activated in your mind. As a result of its activation you more quickly and accurately noticed an inconspicuous berry, mushroom or fish bite. Conversely, if you were focused on berry picking, you might “miss” a mushroom you were not prepared to search for. In this case you dealt with a switching mechanism that operates both by forming a salient focal image against the background and by changing search behaviour.
No matter how the human psyche differs from that of other animals, switching between prey types plays an important role in the activity of all polyphagous predators. For example, in experiments with guppy aquarium fish fed Drosophila (surface‑flying flies) and tubifex (small tubeworms that burrow in the substrate), the fish consumed a disproportionately larger amount of the dominant prey type, because they searched for food either near the bottom or at the surface – where the chance of finding food was higher. We cannot conclude whether the mental image of the sought prey changed in the guppies, but we clearly recorded a change in their foraging behaviour.
An important characteristic of a polyphagous predator capable of switching from one prey to another is its response to changes in relative prey density (Fig. IV-10.1). A Type I predator selectively consumes a particular prey species even when it is rare, and its consumption increases with its prevalence. A Type II predator consumes prey of a given species proportionally to its abundance while it is rare, and after a certain threshold its consumption saturates. The most common, however, is the Type III response (demonstrated in the guppy experiment): a rare prey is consumed disproportionately little, and the predator pays “attention” to it only when its abundance reaches a certain limit. The prevalence of Type III responses among polyphagous predators explains why predators equalize prey numbers, exploiting more common species more strongly and giving “rest” to rare ones.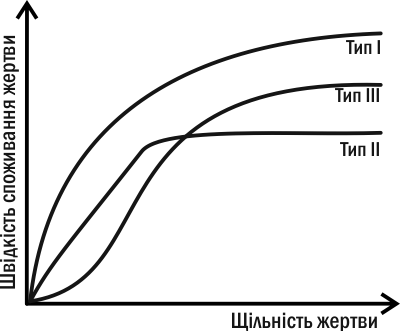
Fig. IV-10.1. Types of polyphagous predator response to changes in the density of one of its prey species
Many morphological and lifestyle features of organisms are explained by their adaptations for defense against predators or for capturing prey. The "arms race" between predators and prey is a major driver of evolution.
For example, plants that have been grazed by animals produce new leaves rich in defensive compounds: phenols, waxes, tannins. When oak leaves are chewed by caterpillars, tannins (polyphenolic compounds stored in specific cell compartments) are released, bind proteins and reduce their nutritional value. Conversely, if trees and shrubs are protected from herbivory, some leaves may become overly shaded by other foliage. In such conditions respiration in the leaves exceeds photosynthesis: this leaf reduces, rather than increases, the overall productivity of the plant. Removal of part of the leaf mass by herbivores actually increases plant productivity!
However, if plant productivity remains constant, herbivores can reach densities that allow them to consume the entire harvest. Many plants (e.g., the cedar pine) exhibit periodic fluctuations in seed production. In a poor‑seed year, few seeds are produced, limiting the populations of seed‑eating mammals and birds. In a mast year, seed abundance is so high that consumers cannot consume it all. This gain for plants comes at a cost: ring growth in trees during mast years declines as much as during complete leaf loss to caterpillars. Synchronization of mast and lean years among trees is driven both by climate and by chemical interactions among plants.
Plant and herbivore adaptations can be extremely tight. One example: on Mauritius the tree Calvaria, or simply the “dodo tree” (Sideroxylon grandiflorum, former name – Calvaria major), was on the brink of extinction because no young trees were present. Biologists realized that Calvaria regeneration ceased about 300 years ago – when the island’s dodos (Raphus cucullatus), large flightless pigeon‑like birds, were extinct. It was hypothesized that Calvaria seeds needed to pass through the dodo’s gut to germinate. To test this, several Calvaria seeds were fed to turkeys. Although some seeds were crushed in the turkey’s stomach, others passed through the digestive tract unharmed and became capable of germination! This insight helped establish artificial restoration of the Calvaria population.
Long‑term prey population responses to predator pressure are not always negative. Predators may eliminate the least valuable individuals in a population (sick, lacking a territory, etc.), or survivors may exhibit compensatory responses. If a predator exploits an over‑crowded prey population, its impact may increase net recruitment or net production.
Typically, predator populations stabilize the numbers of their prey. When prey become scarce, polyphagous predators switch to other prey types, while oligophagous predators reduce their numbers following the decline of their prey due to starvation. Because predators usually reproduce more slowly than their prey, their population recovery is slower than that of the prey. Thus, in typical cases predators cannot completely eradicate their prey!
Under what circumstances can predators wipe out all their prey? Humans have done this repeatedly (e.g., the aforementioned dodos). There is still no consensus among specialists about the role of humans in the extinction of mammoths (Mammuthus primigenius). Likely, the disappearance of this magnificent northern proboscidean resulted from two groups of causes: climate warming and hunting by ancient humans.
Warming climate caused the biome inhabited by mammoths – the so‑called tundra‑steppe or mammoth steppe – to disappear, turning the north into tundra and the south into true steppe. Between tundra and steppe, taiga and broad‑leaf forests intruded. However, it is possible that the scale of ecosystem restructuring was linked not only to climate change but also to human hunting of mammoths and other large herbivores whose grazing maintained the mammoth steppe. Humans also had the potential to eradicate mammoths.
A predator whose numbers depend only weakly on the abundance of a particular prey can completely eliminate that prey. Thus, such a predator must be a polyphage and not specialized on that prey. Humans, with their flexible feeding behavior, could survive even in the absence of mammoths. The danger threatens those prey that remain attractive to predators even at low densities and will be preferentially consumed. This trait characterized the human‑mammoth pair: even when mammoth numbers were low, they remained a primary target for ancient humans, who could efficiently track and harvest them. Complex behavior allowed humans to successfully hunt mammoths even at low densities. Finally, human population growth outpaced mammoth population growth.
Therefore, even without knowing definitively whether humans played a fatal role in mammoth extinction, we can assume they potentially did and certainly amplified the demographic crisis caused by climate warming.
"The immediate threat to a species’ existence is not a ‘consumer’ but a competitor; it is the competitor and only the competitor. When dingoes – initially domestic dogs brought by people and then feral – appeared in ancient Australia, they did not eliminate any of the species that served as their prey, but they wiped out the large marsupial predators that hunted the same animals. Local predators, the marsupial wolf and the marsupial devil, were far stronger than dingoes, yet in hunting skill these ancient, comparatively stupid and slow beasts were outclassed by ‘modern’ mammals.
Dingoes reduced prey populations so much that the hunting methods of their competitors were no longer ‘profitable’, and thus they now survive only on Tasmania, where dingoes never reached" (K. Lorenz, 2001).
Farmers who hunted Tasmanian wolves managed to eradicate this remarkable marsupial predator. The extermination of their prey by hunters can be compared to predation on them (although hunters’ motivation regarding Tasmanian marsupial wolves was more akin to interference competition). To this day, legends persist in Tasmania that some lucky person has encountered this animal on remote parts of the island, but most likely these stories reflect the wishes of nature enthusiasts rather than harsh reality. In any case, in the era of camera traps and molecular‑genetic analyses, no evidence of the marsupial wolf’s existence has been obtained from potentially interesting biomaterial samples.